
Figura 1: Organización anatómica del sistema nervioso.
La función del sistema nervioso periférico (SNP) es conectar el sistema nervioso central (SNC) a los miembros y órganos del cuerpo. El sistema nervioso periférico (SNP) es el encargado de coordinar, integrar y regular nuestros órganos internos por medio de respuestas involuntarias o actos reflejos.
SISTEMA NERVIOSO SOMÁTICO
FUNCION
Su función es la inervación de la musculatura estriada esquelética mediante fibras nerviosas que salen del sistema nervioso central. Esas fibras axónicas van directamente, sin efectuar ningún relevo, hacia las placas mioneurales de los músculos esqueléticos para hacer sinapsis.
CARACTERISTICA
Está compuesto por vías sensitivas y vías motoras. Diversos estímulos como la temperatura, la presión, el dolor, los relacionados con los sentidos (visión, gusto, olfato, audición) o los que provienen de músculos y tendones (propiocepción) ingresan vía aferente (sensitiva) hasta el sistema nervioso central. Esos estímulos dan por resultado una respuesta motora voluntaria, reflejada en contracciones de la musculatura estriada esquelética.
NERVIOS CRANEALES
Se originan a nivel del tronco encefálico, en las proximidades del cuarto ventrículo. Tienen por función la inervación de los órganos de los sentidos, de músculos, glándulas y órganos internos como el corazón y los pulmones.
- Nervio olfatorio o I par craneal: se origina en la mucosa olfatoria, cruza los agujeros de la lámina cribosa del etmoides y termina en el bulbo olfatorio. Es un nervio puramente sensorial y su función es la olfacción.
- Nervio óptico o II par craneal: se origina en las fibras que provienen de la retina, cruza el agujero óptico de la órbita y termina en el quiasma óptico. Es un nervio sensorial y su función en la visión.
- Nervio motor ocular común o III par craneal: es un nervio mixto aunque principalmente motor. La función motora somática permite el movimiento del párpado y determinados movimientos del globo ocular. La actividad motora parasimpática condiciona la acomodación del cristalino y la constricción de la pupila o miosis.
- Nervio patético o IV par craneal: es un nervio mixto aunque principalmente motor, cuya función motora permite el movimiento del globo ocular.
- Nervio trigémino o V par craneal: es un nervio mixto. La porción sensitiva transmite las sensaciones de tacto, dolor, temperatura y propiocepción de la cara. La porción motora inerva los músculos de la masticación
- Nervio motor ocular externo o VI par craneal: es un nervio mixto aunque principalmente motor, cuya función motora permite movimientos del globo ocular.
- Nervio facial o VII par craneal: es un nervio mixto. La porción sensitiva transporta la sensibilidad gustativa de los 2/3 anteriores de la lengua. La porción motora somática inerva la musculatura de la mímica facial. La porción motora parasimpática inerva las glándulas salivales y lagrimales.
- Nervio auditivo o estatoacústico o VIII par craneal: es un nervio mixto, principalmente sensorial. La función principal es transportar los impulsos sensoriales del equilibrio y la audición.
- Nervio glosofaríngeo o IX par craneal: es un nervio mixto. La porción sensorial transporta la sensibilidad gustativa del 1/3 posterior de la lengua. La porción motora somática inerva la musculatura que permita la elevación de la faringe durante la deglución. La porción motora parasimpática inerva la glándula parótida.
- Nervio vago o X par craneal: es un nervio mixto. La función sensorial transporta la sensibilidad de la epiglotis, faringe, así como estímulos que permiten el control de la presión arterial y la función respiratoria. La porción motora somática inerva los músculos de la garganta y cuello permitiendo la deglución, tos y la fonación. La porción motora parasimpática inerva la musculatura lisa de los órganos digestivos, el miocardio y las glándulas del tubo digestivo.
- Nervio espinal o XI par craneal: es un nervio mixto principalmente motor que inerva músculos deglutorios, el músculo trapecio y el músculo esternocleidomastoideo.
- Nervio hipogloso o XII par craneal: inerva la musculatura lingual.

Figura 2: Sistema nerviosos periférico nervios craneales
NERVIOS RAQUÍDEOS
Emergen de la médula espinal, atraviesan los espacios intervertebrales a lo largo de la columna vertebral y se distribuyen por todo el organismo. Los nervios raquídeos o espinales son todos mixtos y ejercen el control de la mayoría de los músculos esqueléticos, de los músculos lisos y de las glándulas. Es por ello que poseen elementos somáticos relacionados con la piel y la musculatura voluntaria, y elementos viscerales que se relacionan con estructuras glandulares, órganos internos y vasos sanguíneos.
A partir de la sustancia gris presente en la médula espinal nacen dos raíces dorsales, una a la derecha y otra a la izquierda, y dos raíces ventrales, una a la derecha y otra a la izquierda. Las raíces dorsales son aferentes y contienen axones sensitivos que captan la información de los receptores sensoriales ubicados en la musculatura esquelética, en la piel, articulaciones, músculos lisos de las vísceras y glándulas para transportarlos hacia la médula espinal. Las raíces dorsales forman un ganglio raquídeo en su recorrido, compuesto de cuerpos neuronales. Las raíces ventrales son eferentes y llevan axones motores somáticos que llegan directamente a los músculos esqueléticos estriados gobernados por el sistema somático. Además, las raíces ventrales también contienen fibras motoras preganglionares del componente simpático y parasimpático (autónomo). Estas raíces ventrales se van a unir, tanto del lado derecho como del izquierdo, con las raíces dorsales sensitivas, por detrás del ganglio raquídeo, dando origen así a cada par de nervios raquídeos. El tronco de estos nervios tiene apenas unos milímetros, ya que se divide enseguida en una rama anterior (ventral), una rama posterior o dorsal, un ramo meníngeo y un ramo comunicante blanco.

Figura 3: Neuroanatomía
Los 31 pares de nervios raquídeos se distribuyen de la siguiente manera:
- 8 pares de nervios cervicales, que se nombran como C1 a C8.
- 12 pares de nervios torácicos, T1 a T12.
- 5 pares de nervios lumbares, L1 a L5.
- 5 pares de nervios sacros, S1 a S5.
- 1 par de nervios coccígeos.

Figura 4: La médula espinal, anatomía.
Las ramas ventrales de los nervios espinales, con excepción de T2 a T12, se conectan entre sí dando lugar a la formación de plexos nerviosos, cuya finalidad es generan nervios más complejos para la inervación de estructuras como el hombro o las extremidades. Luego de esa formación se dividen varias veces antes de llegar a los órganos que deben inervar. Entre los plexos están el cervical, el braquial, el lumbar y el sacro.
- PLEXO CERVICAL
Formado por las ramas ventrales de los nervios cervicales C1, C2, C3 y C4.
FUNCIÓN
Controla la movilidad y la sensibilidad de los músculos de la cabeza, del cuello, de los hombros y de la zona pectoral. También emite ramas profundas (nervios frénicos) para inervar el diafragma.
El plexo se divide en ramas superficiales y ramas profundas. Las ramas superficiales se reúnen sobre el músculo esternocleidomastoideo para formar el PLEXO CERVICAL SUPERFICIAL, exclusivamente sensitivo (Patten; 1995) (Brazis; 2007). Recoge la sensibilidad de parte de la cabeza, el cuello, y la zona alta del tórax, mediante los ramos sensitivos o cutáneos (Kahle; 2008):
- Nervio occipital mayor (C2): se distribuye por la piel de la región craneal posterior.
- Nervio occipital menor o nervio mastoideo (C2): inerva la piel de la región mastoidea, y región lateral craneal.
- Nervio auricular (C2-C3): inerva la piel del pabellón auricular.
- Nervio cutáneo del cuello (C2-C3): inerva la piel de la región supra y subhioidea.
- Nervio supraclavicular (C3-C4): inerva la piel de la parte superolateral del tórax.
- Nervio supraacromial (C3-C4): inerva la piel del muñón del hombro.
- Nervio occipital menor o nervio mastoideo (C2): inerva la piel de la región mastoidea, y región lateral craneal.
- Nervio auricular (C2-C3): inerva la piel del pabellón auricular.
- Nervio cutáneo del cuello (C2-C3): inerva la piel de la región supra y subhioidea.
- Nervio supraclavicular (C3-C4): inerva la piel de la parte superolateral del tórax.
- Nervio supraacromial (C3-C4): inerva la piel del muñón del hombro.
Las ramas profundas del plexo cervical forman el PLEXO CERVICAL PROFUNDO, exclusivamente motor, a excepción del nervio frénico, que contiene en su espesor algunas fibras sensitivas (Mumenthaler; 2004). Se distinguen:
- Ramas mediales: inervan el músculo largo de la cabeza y largo del cuello.
- Ramas laterales: inervan el músculo elevador de la escápula y romboides (C3-C4). Algunas ramas están destinadas a la formación del nervio accesorio (XI par craneal) que inervará al músculo esternocleidomastoideo (C2) que participa en la rotación cervical del cuello y al músculo trapecio (C3-C4), encargado de la elevación de los hombros (Chad; 2006).
- Ramas ascendentes: inervan el músculo recto anterior menor y músculo recto lateral de la cabeza.
- Ramas descendentes: la unión de fibras procedentes de la raíces C1-C2-C3 y el ipogloso forman el asa del hipogloso que inerva los músculos de la región subhioidea (omohioideo, esternotiroideo, esternohioideo, tirohioideo y genihioideo), que participan en la flexión cervical (Campbell; 2005). La raíz C4, y parte de C3 y C5 se unen formando el nervio frénico.
- Ramas laterales: inervan el músculo elevador de la escápula y romboides (C3-C4). Algunas ramas están destinadas a la formación del nervio accesorio (XI par craneal) que inervará al músculo esternocleidomastoideo (C2) que participa en la rotación cervical del cuello y al músculo trapecio (C3-C4), encargado de la elevación de los hombros (Chad; 2006).
- Ramas ascendentes: inervan el músculo recto anterior menor y músculo recto lateral de la cabeza.
- Ramas descendentes: la unión de fibras procedentes de la raíces C1-C2-C3 y el ipogloso forman el asa del hipogloso que inerva los músculos de la región subhioidea (omohioideo, esternotiroideo, esternohioideo, tirohioideo y genihioideo), que participan en la flexión cervical (Campbell; 2005). La raíz C4, y parte de C3 y C5 se unen formando el nervio frénico.
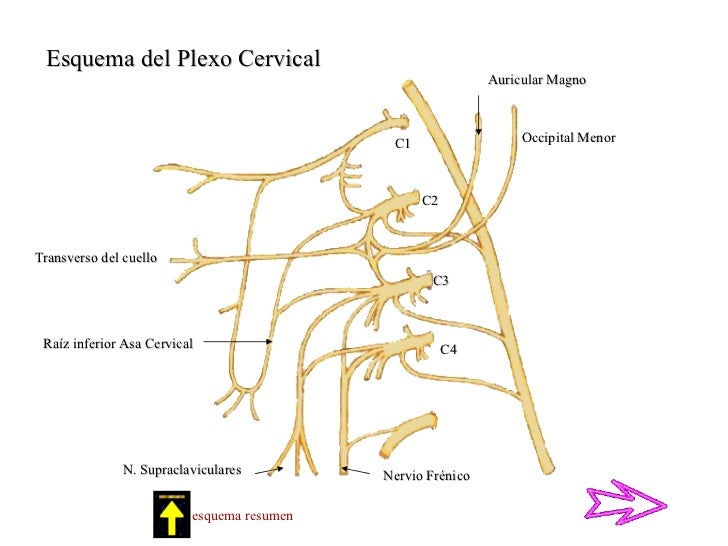
Figura 5:Neuroanatomía de plexos.
- PLEXO BRAQUIAL
El plexo braquial está formado por los ramos anteriores de los nervios espinales de C5, C6, C7, C8 y parte de T1. En su trayecto se distinguen dos porciones: supraclavicular e infraclavicular (Patten; 1995).
A nivel SUPRACLAVICULAR el plexo pasa entre los músculos escaleno anterior y medio. Las fibras nerviosas se distribuyen formando tres troncos primarios:
- Tronco superior: proviene de las ramas anteriores de los nervios C5 y C6 (raramente C4). En él se origina el nervio supraescapular, que va a inervar los músculos supra e infraespinoso, y el nervio subclavio que va a inervar el músculo con el mismo nombre (Chad; 2006).
- Tronco medio: proviene de la rama anterior del nervio C7.
- Tronco inferior: proviene de las ramas anteriores de los nervios C8 y T1.
- Tronco superior: proviene de las ramas anteriores de los nervios C5 y C6 (raramente C4). En él se origina el nervio supraescapular, que va a inervar los músculos supra e infraespinoso, y el nervio subclavio que va a inervar el músculo con el mismo nombre (Chad; 2006).
- Tronco medio: proviene de la rama anterior del nervio C7.
- Tronco inferior: proviene de las ramas anteriores de los nervios C8 y T1.
Una vez sobrepasada la clavícula (región INFRACLAVICULAR) cada tronco se divide en una división anterior y otra posterior, que se van a reorganizar para formar los troncos secundarios, fascículos o cuerdas (Fournier et al; 2009):
- Las divisiones anteriores del tronco superior y medio van a unirse formando el Fascículo o Cuerda lateral. Sus nervios terminales serán: el N. Torácico medial, el N. Cutáneo medial del brazo y del antebrazo, el N. Musculocutáneo, y la rama lateral del N. Mediano.
- La división anterior del tronco inferior formará el Fascículo o Cuerda medial. Sus nervios terminales serán el N. Cubital y la rama medial del N. Mediano.
- Las tres divisiones posteriores formarán el Fascículo o Cuerda posterior. Son ramas terminales el N. subescapular, el N. toracodorsal, el N. axilar y el N. radial (Brazis; 2007) (Kahle; 2008) (Mumenthaler; 2004)
- Las divisiones anteriores del tronco superior y medio van a unirse formando el Fascículo o Cuerda lateral. Sus nervios terminales serán: el N. Torácico medial, el N. Cutáneo medial del brazo y del antebrazo, el N. Musculocutáneo, y la rama lateral del N. Mediano.
- La división anterior del tronco inferior formará el Fascículo o Cuerda medial. Sus nervios terminales serán el N. Cubital y la rama medial del N. Mediano.
- Las tres divisiones posteriores formarán el Fascículo o Cuerda posterior. Son ramas terminales el N. subescapular, el N. toracodorsal, el N. axilar y el N. radial (Brazis; 2007) (Kahle; 2008) (Mumenthaler; 2004)

Figura 6: Neuroanatomía de plexos.
- PLEXO LUMBAR
El plexo lumbar corresponde a las uniones que se establecen entre las ramas anteriores de los nervios raquídeos de L1 a L4, con íntima relación con los fascículos de origen del músculo psoasilíaco. También contribuye a su formación el nervio T12.
- El nervio iliohipogástrico, con fibras de T12 y L1, corre anterior al cuadrado lumbar. Lateral a este perfora el transverso del abdomen donde discurre entre este y el oblicuo interno. Encima de la cresta ilíaca emite un ramo cutáneo lateral, y continua como su ramo cutáneo anterior.
- El nervio ilioinguinal, con fibras de T12 y L1, corre más abajo que el iliohipogástrico. Atraviesa la pared abdominal lateral y entra al conducto inguinal.
- El nervio genitocrural, con fibras de L1 y L2, perfora al músculo psoas mayor para salir por su superficie anterior. Después de pasar posterior al uréter, se divide en sus ramos genital y femoral. El ramo genital entra al conducto inguinal, y el ramo femoral corre por lateral a la arteria ilíaca externa y luego entra en la vaina femoral junto a la arteria femoral.
- El nervio cutáneo femoral lateral, con fibras de L2 y L3, sale por el borde lateral del psoas, y va hacia la espina ilíaca antero-superior. Corre posterior al ligamento inguinal, en la llamada lacuna vasorum y entra en el muslo.
- El nervio obturador, con fibras de L2 a L4, corre detrás del psoas, en su lado medial. Luego pasa por la línea terminal y pasa por la cavidad pélvica, entra al conducto obturador, para llegar al muslo. Acá se divide en sus ramos.
- El nervio femoral, con fibras de L2 a L4, es el más grande de los nervios del plexo lumbar. Sale por el borte inferior lateral del psoas, para correr entre éste y el músculo ilíaco. Luego pasa posterior al ligamento inguinal, para entrar al compartimiento anterior del muslo. Ahí se divide en sus múltiples ramos.

Figura 7: Neuroanatomía de plexos.
- PLEXO SACRO
En anatomía humana, el plexo sacro es una red de nervios constituido por las anastomosis que forman las ramas anteriores de los nervios sacros (L5-S4) y fibras anteriores lumbares (T12-L4), repartiendo inervación a la pelvis y los miembros inferiores.
El plexo sacro está formado específicamente por:
- un tronco (tronco lumbosacro) constituido por ramas anteriores del cuarto nervio lumbar (L4) que se unen con las ramas anteriores de L5;
- fibras de la división anterior de los nervios sacros
Los nervios que así forman el plexo sacro convergen en dirección de la parte baja del agujero ciático mayor y se unen formando una banda aplanada triangular. De las superficies anterior y posterior emergen varias ramas.
La banda misma continúa como el nervio ciático, el cual se divide en la parte posterior del muslo en dos nervios, el nervio tibial y el nervio fibular común. En ocasiones, estos dos nervios emergen por separado del plexo, aunque en todos los casos se vuelve uno independiente del otro.
A menudo, el plexo sacro y el plexo lumbar son considerados un plexo único, llamado en ese caso, el plexo lumbosacro.
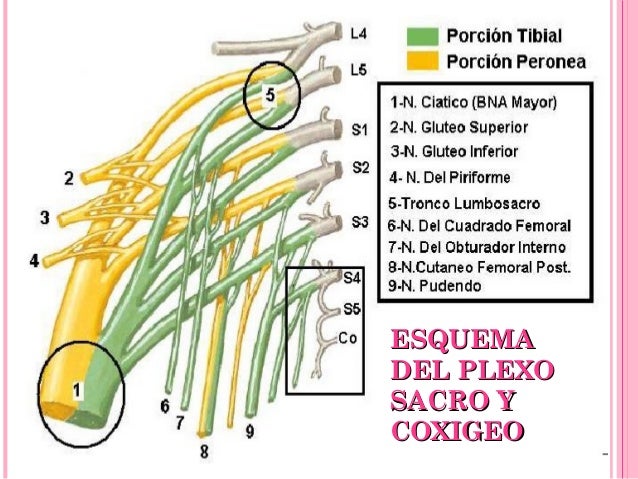
Figura 8: Neuroanatomía de plexos.
- DERMATOMAS
Un dermatoma es el área de piel inervada por un nervio espinal, correspondiente a un segmento medular. (Snell; 1997b)
Los dermatomas se superponen, por lo que para que exista una zona de anestesia completa deben lesionarse más de un nervio espinal. Esta superposición es aún mayor para la sensibilidad termoalgésica que para la táctil, por lo que el área de pérdida de sensibilidad táctil es mayor que el de la termoalgésica para una misma afectación nerviosa espinal.
Los dermatomas se distribuyen de forma horizontal descendente a lo largo del tronco y de forma longitudinal en las extremidades.
El mapa de dermatomas sensitivos nos ayudará en la localización de la lesión medular cuando en la exploración encontramos una hipoestesia o anestesia hasta un nivel sensitivo. Este hallazgo nos indicará que la lesión puede estar en dicho nivel medular o superior, debido a la disposición de las fibras espinotalámicas.

Figura 9: Dermatomas.
- MIOTOMAS
En el desarrollo embrionario de los vertebrados, un miotoma es un grupo de tejidos formado de los somitas. Estos somitas se desarrollan en la pared muscular del cuerpo.
Cada miotoma se divide en una parte epaxial dorsal y una parte hipoaxial ventral.
Los mioblastos de la división hipoaxial forman los músculos de las paredes abdominales y torácicas anteriores. __
El término "miotoma" también se usa para describir los músculos inervados por una raíz nerviosa. Es el equivalente motor al dermatoma.
Cada musculo del cuerpo está inervado por un nivel particular o segmento de la médula espinal y su correspondiente nervio espinal. El músculo y su nervio forman el miotoma.
La masa del músculo epaxial pierde su caracter segmental para formar los músculos extensores del cuello y tronco de los mamíferos. En peces, salamandras, cecílidos y reptiles, la musculatura corporal permanece segmentada como en el embrión, aunque a menudo se pliega y se superpone, con masas epaxiales e hipoaxiales divididas en varios grupos musculares distintos.
En los humanos, las pruebas de los miotoma pueden ser una parte integral de la examinación neurológica ya que cada raíz nerviosa que sale de la médula espinal suple a un grupo específico de músculos. Las pruebas de miotomas, en forma de pruebas musculares isométricas resistidas, proporciona al médico información del nivel donde se encuentra la lesión en la médula.
Durante la prueba de los miotomas, el médico mide la debilidad muscular de un grupo particular de músculos. el resultado puede indicar la lesión de la raíz nerviosa de la médula espinal, o una herniación del disco intervertebral en las raíces nerviosas espinales.
La distribución de los miotomas de las extremidades superiores e inferiores son de la siguiente manera:
- C1/C2-flexión/extensión del cuello
- C3-Flexión lateral del cuello
- C4-Elevación del hombro
- C5-Abducción del hombro
- C6-Flexión del codo/extensión de la muñeca
- C7-Extensión del codo/flexión de la muñeca
- C8-extensión del pulgar
- T1-Abducción del dedo
- L2-Flexión de la cadera
- L3-Extensión de la rodilla
- L4-Dorsi-flexión del tobillo
- L5-Extensión del dedo gordo
- S1-Flexión plantar del tobillo
- S2-Flexión de la rodilla

Figura 10: Miotomas.
SISTEMA SENSORIAL
La selección natural ha diseñado mecanismos biológicos sensibles a diferentes tipos de energía relevante para la supervivencia y la reproducción, son los sistemas sensoriales, también llamado sentidos.
Los sentidos son vías por las que la información acerca de parte de la realidad circundante accede y al SN de las especies que lo poseen. Cualquier tipo de estimulación que pueda influir sobre la conducta, para poder hacerlo, tiene que haber sido transformada en cambios eléctricos neuronales, especialmente en impulsos nerviosos.
FISIOLOGÍA DE LOS SENTIDOS
Los diferentes tipos de energía susceptibles de influir sobre la conducta de un organismo reciben el nombre genérico de estimulación sensorial. La vía a través de la cual los estímulos llegan al organismo, esencialmente al SNC y rigen la conducta (o influyen sobre ella) está constituida por los receptores sensoriales localizados en los órganos de los sentidos.
1. Tipos de energía estimular: la modalidad sensorial La estimulación sensorial constituye el aspecto físico y material de los estímulos, su modalidad (luz, sonido…) y sus características físicas de frecuencia, amplitud o intensidad… No todas las energías afectan al comportamiento ni todas forman parte del acervo de posibles estimulaciones.
Además de detectar y “clasificar” la energía, (extraer la información relativa al tipo de energía), el SN debe ser capaz de codificar su procedencia, intensidad y duración. A la transformación de las diferentes modalidades energéticas en actividad eléctrica llevada a cabo por los receptores sensoriales se le llama transducción sensorial.
El problema al que se enfrente el SN a la hora de proporcionar información sobre las diferentes modalidades sensoriales es que todas ellas son codificadas en términos eléctricos a nivel neuronal.
2. La cuantificación de la energía estimular: codificación de la intensidad (o amplitud) del estímulo
Las neuronas sensoriales “informan” de la intensidad de la energía incidente, ello hace posible que podamos afirmar que una sensación es más intensa que otra.
2 son los mecanismos con los que cuentan las neuronas sensoriales:
- código de frecuencia: consistente en que a medida que aumenta la intensidad de la estimulación, aumenta correlativamente la frecuencia de potenciales de acción que una neurona sensorial transmite. Una neurona no puede generar más de unos 1.000 potenciales de acción por segundo. Cuando una determinada intensidad de estimulación le haga responder con esa frecuencia de disparo, cualquier aumento de esa intensidad pasará desapercibido para esa neurona.
Hay una estrategia que permite ampliar el rango de intensidades codificables sirviéndose de una característica de las neuronas sensoriales: umbral de respuesta. Cada neurona empieza a responder sólo cuando la estimulación tiene la intensidad necesaria. Este umbral difiere de unas neuronas a otras y mientras que unas responden a estimulaciones muy débiles (son muy sensibles) otras sólo lo hacen cuando la estimulación es intensa o muy intensa.
3. La codificación de la duración del estímulo Un fenómeno bastante habitual es el de la disminución o incluso desaparición de la sensación aun cuando la estimulación sensorial sigue teniendo la misma intensidad. Cuando esta reducción se explica por la fisiología de los receptores sensoriales recibe el nombre de adaptación sensorial, mientras que cuando los receptores siguen respondiendo de la misma forma, pero el individuo deja de respondes a la estimulación, se suele hablar de habituación, que es un tipo de aprendizaje no asociativo cuya explicación hay que buscarla en cambios fisiológicos en las sinapsis entre neuronas dentro del SNC, o en procesos más complejos.
Hay unos receptores que son más fidedignos que tros, tanto en lo que se refiere a la codificación de la intensidad de la estimulación como a su duración. El caso más característico es el de los receptores de presión táctil asociados a los corpúsculos de Pacini receptores (son neuronas) que sólo responden con potenciales de acción al principio y al final de la estimulación y dejan de hacerlo el resto del tiempo, lo que significa que responden a los cambios en el estímulo. Esta reducción es la adaptación sensorial.
El proceso de adaptación sensorial no sucede de la misma forma en todos los receptores sensoriales, algunos se adaptan muy rápidamente, otros tardan mucho en adaptarse o nos e adaptan en absoluto (como es el caso de algunos receptores del dolor). Los receptores que se adaptan deprisa reciben el nombre de receptores fásicos y los que tardan mucho o no son adaptables se les denomina receptores tónicos.
4. La localización del estímulo: campos receptivos e inhibición lateral
La ubicación o localización del estímulo se basa en la distribución estructural de los receptores sensoriales, en el modo en que se relacionan los receptores sensoriales con las diferentes zonas centrales de procesamiento (SNC) de la información que recogen. Esto se explica por la diferente densidad de receptores en unas zonas que en otra. La actividad fisiológica de una neurona sensorial resulta modificada por la estimulación de los receptores que contactan con ella, es lo que se conoce como campo receptivo.
La interacción entre campos receptivos próximos explica el fenómeno denominado inhibición lateral. Es una observación bien contrastada que percibimos mejor los cambios en la intensidad de la estimulación sensorial o zonas de contraste. La estimulación de un campo receptivo no sólo excita a la neurona sensoria, sino que inhibe a las que tienen campos receptivos próximos.
LOS RECEPTORES SENSORIALES Y LA TRANSDUCCIÓN
El estímulo produce cambios en el potencial de membrana de los receptores sensoriales que son de tipo graduado similares a los potenciales postsinàpticos. Este cambio de potencial recibe el nombre de potencial de receptor (o potencial generador) si el receptor sensorial es una neurona. Este potencial de receptor es una despolarización, aunque los potenciales originados en los receptores visuales son una excepción, pues son hiperpolarizados.
En todas las modalidades sensoriales se han de generar potenciales de receptor que determinaran, bien directa o indirectamente el disparo de potenciales de acción en las neuronas sensoriales. Para que un organismo pueda verse afectado por una estimulación ambiental, es preciso que el cambio de potencial sobrepase el umbral de disparo de la neurona sensorial y se transforme en un potencial de acción.
A. Quimiorreceptores y transducción
1. Receptores gustativos
Los botones gustativos no se distribuyen de modo uniforme por la lengua, sino que se concentran en las papilas gustativas.
Las células gustativas proyectan extensiones en forma de microvellosidades hasta el poro en que termina el botón gustativo, mientras que en el otro extremo hacen sinapsis con las neuronas sensoriales. Existen 4 cualidades gustativas: dulce, salado, amargo y ácido y se añade el glutamato monosódico, el umami.
Transducción gustativa: los receptores gustativos funcionan de forma similar a las neuronas en la medida en que las sustancias estimulantes actúan sobre receptores de membrana, bien abriendo canales iónicos, bien activando receptores proteínicos que inician una cascada metabólica responsable de la activación de segundos mensajeros (según el tipo de sabor, el mecanismo será uno u otro). En ambos casos el resultado es una despolarización subsecuente a la apertura o cierre de los canales de Na+, K+ o Ca2+ que es de tipo graduado, produciéndose la liberación de un neurotransmisor químico en la sinapsis entre el receptor y la neurona sensorial.

Figura 11: Botón Gustativo.
2. Receptores olfatorios
Feromonas: son unas sustancias olorosas específicas, capaces de señalar y provocar cambios fisiológicos en los congéneres.
Las sustancias olorosas lo son porque algunas de sus moléculas se esparcen por el aire y entran en contacto con los receptores olfativos, neuronas receptoras olfatorias o receptores olfatorios. En el hombre estos receptores se encuentran en la mucosa olfatoria. Esta mucosa consta de células receptoras (CR), células de soporte (CS) y células basales (CB).

Figura 12: Cavidad nasal.
Los receptores olfatorios proyectan cilios (ramificaciones finas de la única dendrita que poseen) hacia el interior de la capa mucosa. Los receptores olfatorios son neuronas bipolares con una proyección dendrítica hacia la superficie de la mucosa olfatoria donde se engrosa y proyecta sus cilios y una proyección axónica en su polo opuesto que atravesando el hueso craneal a través de la placa cribiforme conecta ipsilateralmente con el bulbo olfatorio.

Figura 12: Organización celular de la mucosa olfatoria.
La transducción olfativa se inicia en los cilios de las neuronas receptoras, ya que si éstos se eliminan, queda abolida la respuesta olfativa. La frecuencia de los potenciales de acción depende de la concentración de la sustancia olorosa, pero una exposición prolongada da lugar a una reducción en la frecuencia de los potenciales hasta niveles relativamente bajos, esto se conoce como adaptación sensorial.
3. Receptores nociceptivos
Transducción de la información nociceptiva: hay determinados procesos químicos que tienen lugar en los tejidos y afectan a los receptores sensoriales que informan sobre el dolor.
Hay muchos compuestos tanto exógenos como endógenos que afectan a las terminaciones sensoriales nociceptivas. Algunas de estas sustancias activan directamente a las neuronas sensoriales y otras intervienen en los procesos de nocicepción actuando como moduladores modificando el umbral de respuesta de las terminaciones libres a otros estímulos químicos o mecánicos. Diferentes agentes químicos sensibilizan o estimulan las terminaciones nerviosas libres produciendo dolor. Los tejidos lesionados y las células del sistema inmunitario liberan diversos compuestos que excitan a los nociceptores. Además, la estimulación de los nociceptores puede producir la liberación de sustancias que ellos mismos sintetizan.
B. Mecanorreceptores y transducción.
1. Receptores cutáneos
Los Mecanorreceptores responden a la acción de fuerzas mecánicas. Entre ellos se encuentran parte de los receptores sensoriales de la piel, y los de los músculos, los tendones y las articulaciones. Los Corpúsculos de Pacini son receptores que responden a la deformación mecánica de la piel de forma que la presión ejercida por el estímulo genera cambios en la conformación de las láminas del tejido conectivo que rodean la terminación sensorial del receptor, al igual que sucede cuando esta presión se retira. Estos receptores son neuronas cuya ramificación periférica es la fibra aferente que actúa como receptor sensorial, mientras que su ramificación central responde con potenciales de acción transmitiendo la información al SNC. Los cuerpos celulares de estas neuronas se encuentran en los ganglios de la raíz dorsal y en los ganglios encefálicos. Su estructura le hace especialmente sensible para señalar los cambios rápidos de presión.

Figura 13: Corpúsculos de Pacini.
La transducción es la transformación de la energía mecánica en potenciales generadores por parte de los corpúsculos de Pacini se lleva a cabo de la siguiente forma:
- el estímulo que incide sobre la piel es modificado por las células accesorias que rodean al receptor
- el estímulo mecánico incide sobre la membrana de la zona receptora de la neurona sensorial para producir un cambio en su potencial de membrana
- este cambio de potencial puede producir un potencial de acción en la fibra nerviosa aferente.
2. Receptores Auditivos
La transducción auditiva es el resultado del efecto de las ondas sonoras (fruto de las oscilaciones en la concentración de moléculas del aire que resultan de cualquier proceso vibratorio) que son una forma de fuerza mecánica ejercida por las moléculas del aire, sobre os receptores auditivos: las células ciliadas.
La onda sonora, en su recorrido hasta la cóclea tiene que pasar por el meato auditivo. En su desplazamiento hacia el interior del oído, la onda sonora “tropieza” con la membrana timpánica. Las variaciones de presión de la onda sonora se transmiten al tímpano que empieza a vibrar con una frecuencia y una amplitud proporcional a la frecuencia e intensidad de la onda sonora que llega. El tímpano y la cadena de huesecillos del oído medio constituyen una adaptación específica para captar ondas sonoras que viajan a través del aire: originariamente, el oído era una estructura especializada en recoger las vibraciones del medio líquido.
Las vibraciones del aire no son capaces por sí solas de afectar al líquido coclear de los animales terrestres, ya que las ondas sonoras que discurren por el aire apenas afectan a los medios líquidos, sino que se reflejan casi por complejo en ellos. El tímpano y la cadena de huesecillos que conectan el efecto de la onda o vibración sonora, a través de la ventana oval, con el almacén de líquido que es la cóclea constituyen el sistema de amplificación que permite que las ondas sonoras de tipo aéreo se transmitan a un medio líquido sin apenas pérdida.
El sistema de amplificación se consigue gracias a la interacción de 2 factores:
- la cadena de huesecillos actúa como un sistema de palancas que hace que la vibración del tímpano aumente su fuerza y reduzca su amplitud al llegar a la ventana oval.
- (el más importante) el hecho de que el tímpano tenga una superficie de alrededor de 70mm cuadrados y la ventana oval 3mm cuadrados, lo que determina que la fuerza de la vibración de ésta se vea multiplicada.
Cuando las ondas sonoras llegan a la ventana oval sus vibraciones se transmiten a la perilinfa que rellena la rampa vestibular de la cóclea. Como el líquido coclear no se puede comprimir, cada vez que la ventana oval oscila hacia dentro de la cóclea aumenta la presión del líquido, mientras que cuando oscila hacia fuera, se reduce dicha presión esto significa que la onda sonora se ha transformado en la cóclea en una onda de presión que recorre la perilinfa líquida.
La vibración de la perilinfa afecta a las 2 membranas que delimitan la rampa media o conducto coclear, relleno de endolinfa. Estas membranas son:
- la membrana de reissner o vestibular por arriba
- la membrana basilar por abajo.
Ambas membranas sufren una deflección, concretamente la de la membrana basilar.

Figura 14: Estructuras esenciales del oído.
C. Fotorreceptores y transducción visual
El procesamiento de la información visual se inicia con la transformación de la luz en señales eléctricas por parte de los fotorreceptores de la retina del ojo. Los fotorreceptores no producen potenciales de acción tras ser estimulados por la luz, sino que responden con cambios en su potencial de membrana (potenciales locales).

Figura 15: Componentes del ojo.
Cuando llega la luz la córnea y el cristalino la enfocan y tras atravesar el humor vítreo, es absorbida por las células fotorreceptoras de la retina. Estas células se disponen sobre el epitelio pigmentado que abarca todo el fondo del ojo y contiene grandes cantidades de melanina, cuya función es recoger la luz que las células retinianas no han podido absorber. Ello impide que haya un reflejo de la imagen que entra desde el fondo del ojo hacia la retina, ya que podría distorsionarla.
Para permitir que la luz llegue a las células fotorreceptoras sin ser dispersada o absorbida las capas celulares de la retina más próximas al cristalino no tienen mielina y son relativamente transparentes.
La retina tiene una pequeña invaginación en forma de mella, situada aproximadamente, en su punto central denominada fóvea. En la fóvea, los cuerpos neuronales de las células retinianas se disponen desplazados hacia los laterales para dejar paso libre a la luz para que no haya distorsión y la luz llegue con menos dificultad. En la parte central de la fóvea, llamada foveola, prácticamente sólo hay células fotorreceptoras en alta concentración que permite que la agudeza visual sea máxima en esa zona.
Hay 2 tipos de células fotorreceptoras llamadas conos y bastones. Hay más cantidad de bastones que conos. En ambos fotorreceptores existen moléculas sensibles a la luz llamadas pigmentos visuales o fotopigmentos. Los bastones contienen un tipo de pigmento específico, mientras que hay 3 tipos de pigmentos en los conos que determinan 3 tipos de conos según el pigmento que usan.

Figura 16: Organización de la retina.
El pigmento visual de los bastones es más sensible a la luz que los de los conos, puesto que les permite captar más luz, y además, los bastones amplifican más la señal luminosa que los conos. Un único fotón1 puede producir una señal eléctrica detectable en os bastones, mientras que se precisan centenares de fotones para que un cono pueda generar la misma respuesta. Por lo tanto, los bastones participan en la visión nocturna o con luz tenue, mientras que los conos se activan durante la visión diurna. Tanto los conos como los bastones hacen sinapsis con neuronas bipolares.
LA TRANSMISIÓN DE LA INFORMACIÓN SENSORIAL AL SISTEMA NERVIOSO CENTRAL
El resultado de la transducción es la generación de potenciales de receptor, que finalmente pueden provocar el disparo de potenciales de acción en las neuronas sensoriales, de forma directa o indirecta, según sea el receptor una neurona o una célula especializada no nerviosa. Los axones de estas neuronas sensoriales constituyen los nervios espinales y craneales y a través de ellos la información es transmitida a diferentes divisiones del SNC.
1. Aproximación General a los Circuitos Sensoriales del SNC
Una de las características fundamentales del procesamiento de la información sensorial en el SNC es que se requiere la intervención de diversos núcleos y regiones encefálicas que funcionan de forma integrada. Existe una especialización en las funciones que desempeña cada una de estas regiones encefálicas, de modo que las señales sensoriales fluyen desde niveles inferiores a niveles superiores, estableciéndose una jerarquía en el procesamiento sensorial.
Esta organización jerárquica implica que la información procedente de los receptores sensoriales llega principalmente a la médula espinal y a regiones situadas en el tronco del encéfalo y en el diencéfalo para alcanzar finalmente la corteza cerebral (excepto la información olfativa que llega en primer lugar a los hemisferios cerebrales).
.
SISTEMA NERVIOSO AUTÓNOMO
El sistema nervioso autónomo (SNA) o vegetativo es la parte del sistema nervioso central y periférico que se encarga de la regulación de las funciones involuntarias del organismo, del mantenimiento de la homeostasis interna y de las respuestas de adaptación ante las variaciones del medio externo e interno. Así pues, ayuda a controlar, entre otras funciones, la presión arterial, la motilidad y secreciones digestivas, la emisión urinaria, la sudoración y la temperatura corporal. Algunas de estas funciones están controladas totalmente por el sistema nervioso autónomo, mientras que otras lo están parcialmente.
- ANATOMÍA DEL SISTEMA NERVIOSO AUTÓNOMO
Sistema nervioso autónomo central
No existe un centro bien definido puramente central del sistema nervioso autónomo. La integración de las actividades del sistema nervioso autónomo ocurre a todos los niveles del eje cerebroespinal y la actividad eferente puede ser iniciada a partir de centros localizados en la médula espinal, tronco encefálico e hipotálamo. La corteza cerebral es el nivel más alto de integración somática y del sistema nervioso autónomo. El sistema nervioso autónomo habitualmente regula las funciones de los órganos mediante reflejos viscerales inconscientes y que en ocasiones se producen como respuesta a cambios en actividades somáticas motoras y sensoriales. Aunque la mayoría de las funciones reguladas por el sistema nervioso autónomo se encuentran fuera del control consciente, las emociones y los estímulos somatosensoriales lo pueden influenciar profundamente. En el sistema nervioso central, las regiones reguladoras sensoriales y autonómicas responden a menudo al mismo tipo de estímulo somático o visceral, de forma que un mismo estímulo es capaz de desencadenar respuestas autonómicas, antinociceptivas y de comportamiento.

Figura 17: Sitema nervioso periferico (Ramon Pignatelli)
Sistema nervioso simpático
Los nervios simpáticos tienen origen en la médula espinal entre los segmentos T-1 y L-2 y desde aquí se dirigen a la cadena simpática paravertebral y finalmente a los tejidos y órganos periféricos. El cuerpo celular de las fibras preganglionares se localizan en el cuerno intermedio-lateral de la médula espinal, que abandonan a través de la raíz anterior junto con las fibras motoras; las fibras simpáticas preganglionares abandonan el nervio espinal inmediatamente después de que éste salga por el agujero de conjunción y constituyen las ramas comunicantes blancas, mielinizadas, que se dirigen hacia la cadena simpática paravertebral.
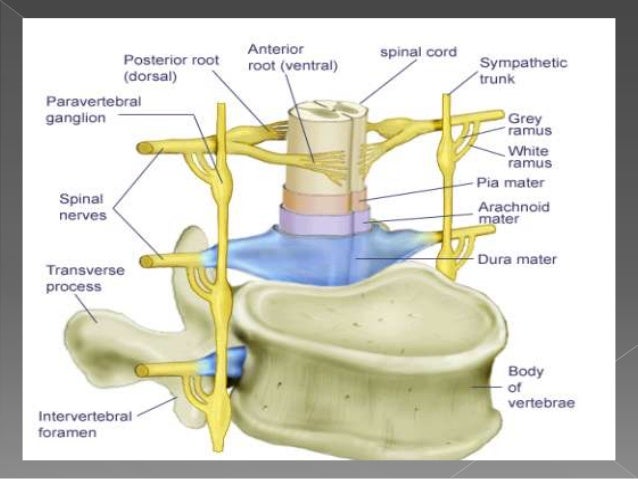
Figura 18: Anatomía del sistema nervioso simpático
Las fibras simpáticas originadas en T-1 generalmente siguen la cadena simpática hacia la cabeza y las de T-2 van hacia el cuello. De T-3 a T-6 se distribuyen al tórax, de T-7 a T-11 al abdomen y de T-12 a L-2 a las extremidades inferiores. Ésta es una distribución aproximada y siempre se dan superposiciones. Prácticamente todos los órganos reciben inervación simpática y la distribución de los nervios simpáticos para cada órgano va a depender de la posición en la que éste se encuentra originariamente en el embrión (por ej. el corazón recibe inervación procedente de la cadena simpática cervical, ya que es en el cuello donde tiene su origen embrionario). La cadena simpática cervical está constituida por fibras procedentes de T1 a T5 que dan lugar a tres ganglios cervicales: superior, medio y cérvico-torácico.
Sistema nervioso parasimpático
Las fibras nerviosas parasimpáticas tienen origen en el tronco encefálico, en los núcleos de los pares craneales III (oculomotor), VII (facial), IX (glosofaríngeo) y X (vago) y en la médula sacra: segundo y tercero nervios sacros, y a veces también del primero y cuarto. El nervio vago tiene la distribución más amplia de todo el SNP, siendo responsable de más del 75% de la actividad parasimpática; inerva al corazón, pulmones, esófago, estómago, intestino delgado, mitad proximal del colon, hígado, vesícula biliar, páncreas y parte alta de los uréteres. En la pared de estos órganos se localiza la neurona postganglionar.

Figura 19: Anatomía del sistema nervioso parasimpático.
Las fibras del III par craneal van a los esfínteres pupilares y músculos ciliares del ojo. Las del VII par inervan a las glándulas lacrimales, sub-maxilares y de la mucosa nasal y las del IX par van hasta la parótida. En estos casos, la neurona postganglionar se localiza en los ganglios de los pares craneales. Las fibras sacras, procedentes sobretodo del segundo y tercer nervios sacros y a veces también del primero y cuarto, se reúnen para formar los nervios pélvicos que se distribuyen por el colon descendente, recto, vejiga, porción baja de los uréteres y genitales externos. La relación de fibras pre y postganglionares es de 1:1 o 1:3, de tal forma que una neurona preganglionar forma sinapsis con muy pocas neuronas postganglionares, lo que asociado a la 7 proximidad de la sinapsis al órgano inervado, lleva a que la estimulación parasimpático sea más localizada, al contrario de lo que sucede en el SNS.
FUNCIONES DEL SISTEMA NERVIOSO AUTÓNOMO
Tono simpático y parasimpático
Tanto el sistema nervioso simpático como el sistema nervioso parasimpático mantienen una actividad constante que se conoce como tono simpático y tono parasimpático. Ello permite que en un momento dado, la actividad de cada uno de estos dos sistemas pueda aumentar o disminuir. El tono simpático mantiene en condiciones normales a casi todos los vasos sanguíneos constreñidos hasta aproximadamente la mitad de su diámetro máximo. Si aumenta la actividad simpática se producirá una constricción adicional, y en cambio si hay una disminución de la actividad simpática se producirá una vasodilatación. Si no existiera tono simpático sólo podría producirse vasoconstricción. 15 El tono simpático y parasimpático depende de la actividad de los centros superiores del tronco cerebral; una parte del tono simpático resulta de la secreción basal de NA y A por la médula suprarrenal.
Activación del sistema nervioso simpático
El sistema nervioso simpático puede activarse de forma masiva; ésto ocurre cuando el hipotálamo es activado por un determinado evento, como un estímulo doloroso o emocional muy intensos y en consecuencia se produce una reacción generalizada en todo el organismo, conocida como reacción de alarma o de estrés, también denominada reacción de lucha o huída (“fight or flight”). Con la descarga simpática masiva el organismo se pone en marcha para llevar a cabo una actividad muscular enérgica; aumenta la presión arterial, aumenta el flujo sanguíneo en los músculos activos y lo disminuye en los órganos innecesarios para una actividad rápida, aumenta de forma generalizada el metabolismo celular, la glucólisis muscular, la fuerza muscular, y la actividad mental. Todo ello permite realizar una actividad física agotadora.
Activación del sistema nervioso parasimpático
Al contrario de lo que ocurre con el sistema nervioso simpático, el sistema nervioso parasimpático se relaciona con procesos de descanso (“rest and digest”), y su activación esta orientada al ahorro de energía. Así, la activación parasimpática produce una disminución de la frecuencia cardiaca, de la velocidad de conducción seno-auricular y aurículo-ventricular, constricción del músculo liso bronquial, miosis, etc. Los signos de descarga parasimpática son las náuseas, vómitos, aumento del peristaltismo intestinal, enuresis y defecación. También origina un aumento de las secreciones. Sin embargo, la mayor parte de los reflejos parasimpáticos son relativamente específicos; así por ejemplo, es posible una bradicardia por estímulo vagal, sin otras alteraciones concomitantes ya que los reflejos parasimpáticos cardiovasculares suelen actuar sólo sobre el corazón; otro ejemplo es el reflejo de vaciamiento rectal, sin afectación del resto del intestino. A veces los reflejos parasimpáticos pueden asociarse, como en el caso del vaciamiento vesical y rectal.
INERVACIÓN AUTONÓMA
Corazón
El corazón recibe abundante inervación simpática y parasimpática que regulan fundamentalmente la frecuencia cardiaca (cronotropismo) y la contractilidad (inotropismo). Las fibras parasimpáticas vagales se dirigen hacia el ganglio estrellado y a partir de aquí acompañan a las fibras simpáticas eferentes cardíacas constituyendo el plexo cardíaco, que es mixto y formado por fibras simpáticas y parasimpáticas. Las fibras parasimpáticas se distribuyen principalmente al nodo sinusal, aurículoventricular y en menor grado a la aurícula, con muy poca o nula distribución ventricular; su efecto principal es el cronotrópico negativo (disminución de la frecuencia cardiaca por disminución de la descarga del nodo sinoauricular y disminución de la velocidad de conducción auriculoventricular). El sistema nervioso simpático tiene la misma distribución supraventricular que el sistema nervioso parasimpático, pero con una distribución ventricular mucho más importante y su efecto predominante es sobre el inotropismo. El tono simpático normal mantiene la contractilidad un 20% por encima de la que habría en ausencia de estímulo simpático. El flujo sanguíneo coronario está regulado principalmente por factores locales relacionados con los requerimientos metabólicos del miocardio, y clásicamente se atribuye poca influencia del sistema nervioso autónomo en la circulación coronaria; sin embargo actualmente existen evidencias de la influencia del sistema nervioso simpático sobre la regulación de los pequeños vasos de resistencia y los grandes vasos de conductancia provocando vasoconstricción.

Figura 20: Inervación autónoma del corazón.
Circulación periférica
El sistema nervioso simpático es sin lugar a duda, el sistema de regulación más importante en la circulación periférica. El sistema nervioso parasimpático ejerce un efecto mínimo en la circulación periférica. El tono vasomotor se mantiene por la acción constante del sistema nervioso simpático, a partir del centro vasomotor del tronco encefálico. La adrenalina de la médula suprarrenal tiene un efecto aditivo. Este tono mantiene a las arteriolas y las vénulas en un estado de constricción parcial, con un diámetro intermedio, con la posibilidad de vasoconstricción adicional o por el contrario de vasodilatación; si el tono basal no existiera, el sistema nervioso simpático solo podría ejercer un efecto vasoconstrictor sin posibilidad de vasodilatación arteriolar. Los cambios en la constricción arterial se manifiestan como cambios de resistencia al flujo sanguíneo. En cambio el sistema venoso es un sistema de capacitancia y no de resistencia y el tono venoso produce una resistencia al flujo mucho menor que en el sistema arterial y los efectos de la estimulación simpática alteran la capacidad más que la resistencia del sistema venoso. Como el sistema venoso funciona como un reservorio de aproximadamente el 80% del volumen sanguíneo, pequeños cambios en la capacitancia venosa producen grandes cambios en el retorno venoso y por tanto en la precarga cardiaca.
Sistema respiratorio
La inervación simpática llega a través de las fibras postganglionares del ganglio estrellado y la parasimpática proviene del nervio vago. Las fibras simpáticas y parasimpáticas viajan juntas al igual que sucede en el corazón e inervan el músculo liso bronquial y vascular pulmonar. La estimulación simpática produce broncodilatación y también vasoconstricción pulmonar si bien que los factores locales parecen ejercer un efecto mucho más importante en la regulación de la circulación pulmonar (vasoconstricción pulmonar hipóxica). El sistema nervioso parasimpático, por el contrario, es broncoconstrictor y además aumenta las secreciones bronquiales.
REFERENCIAS
- Gustavo Ramon S; conocimiento corporal II
- col.egi oficial infermeres I infermes. 2001
- Anatomía del sistema nervioso- cap 3
- . Guyton AC, Hall JE. El sistema nervioso autónomo; la médula suprarrenal. En: Tratado de Fisiología Médica. Madrid: McGraw-Hill Interamericana de España; 1996. p. 835-847.
- Gershon MD.The Second Brain. Nueva York: Harper Collins; 1998.
- -Chad D (2006). Enfermedades de las raíces y plexos nerviosos. En: Bradley PW (Ed), Neurología clínica, (pp 2247-2275). Madrid: Elsevier.
- Kahle W (Ed) (2008). Médula espinal y nervios espinales. En: Atlas de Anatomía con correlación clínica, (pp 48-97). Madrid: Panamerican
- Antigüedad A, Zarranz JJ, Barcena J (1990). Recuerdo anatómico y síndromes topográficos del Sistema Nervioso Central. En: Medicine. 1(61): 2389-2419.
- Snell SR (Ed) (1997b). Dermatomas y actividad muscular. En: Neuroanatomía Clínica, (pp 138-150). Madrid: Editorial Médica Panamericana.
- Ciencias biologicas y educacion para la salud (2010).
- Psicobiologia (2011)
Hecho por Wendy Gonzalez


No hay comentarios.:
Publicar un comentario